
The soil market: plants spend some of their carbon on buying nutrients from mycorrhizae
Similar to the way that we take up carbon (C) through the food we eat, plants absorb C in the form of CO2 from the atmosphere through photosynthesis, and use it to build sugars (Fig. 1). After having a good snack, our body decides where to send the C it has just gained: should I use it to grow muscles (or fat), or do I need to burn it in order to work out, now? If we burn this C, we breathe it out as CO2 in respiration. Similarly, plants also decide how to allocate their sugary C, and their ‘duties’ also include growth and respiration.
Whilst we get all the building blocks that we need to grow and live from food, plants have to ‘shop for’ nutrients, such as nitrogen and phosphorus in soil, and this also costs them C. However, nutrients are sometimes unavailable in local shops. One way plants then obtain the less available nutrients is by making long-term deals with so-called mycorrhizal fungi (mycorrhizae) in the soil. Again, the currency for this transaction is C.

Fig. 1: How plants spend their carbon, gained only through photosynthesis: partitioning to plant growth, autotrophic respiration (Raut) and mycorrhizae. Figure credit (right): Sara Vicca, photo credit (left): Melanie Verlinden
Different ecosystems = different budgets, but why?
The fraction of C allocated to different processes is referred to as C partitioning. This is expressed as a fraction of gross primary production (photosynthesis), and is a key process that affects the growth of individual plants, as well as whole terrestrial ecosystems. It determines how much of the CO2 taken up by plants will be released back into the atmosphere as CO2, as well as where and how much of it will be sequestered and stored in the ecosystem. This is important for the pathways through which plant C reaches the soil (e.g. as litter, dead roots, root exudates, mycorrhizal biomass) which can strongly influence its fate. Despite its central role in terrestrial C cycling, C partitioning and its relationship with nutrient availability (i.e., the availability and cost of nutrients in the local shops), remains poorly understood. In some ecosystems, it has been shown that only 40% of the photosynthesis ‘budget’ is invested in plant growth, whereas elsewhere it can be up to 70% [1]. Why this variation occurs is still unknown, but evidence is growing that nutrient availability plays a key role here, and business done with mycorrhizae might be a critical determinant of the price of nutrients.
And that is where our research came in. We added some phosphorus to some corn and measured a bunch of things to quantify C partitioning to unravel the role of mycorrhizae in determining the price of nutrients.
We conducted two nutrient manipulation experiments using mycorrhizae in symbiosis with corn (Zea mays) (Fig. 2, 3) with the aim to quantify C partitioning to all duties (Fig. 4). When phosphorus availability for plants is low (less phosphorus in the local shops), we expected plants to invest more of their C budget relative to the amount of photosynthesis to acquire phosphorus, and to depend on the mycorrhizal fungi to get it. This would leave a smaller fraction of the budget for growing plant biomass. To find out whether this long term business with mycorrhizae was profitable for plants, and whether this changed with nutrient availability, we set up a treatment where soil was sterilised to compare with what happens when there is no mycorrhizae (Fig. 3).

Fig. 2: Scientist are busy measuring C-fluxes in the greenhouse where corn plants are grown under different nutrient and mycorrhizal conditions.

Fig. 3: The set-up of the second experiment (of 2017) with six different nutrient- and mycorrhizae treatments. The green line shows the amount of phosphorus (the thicker the line, the more phosphorus we added, also marked with “P1” to “P4”). “AMF” means plants were inoculated with mycorrhizae while “no_AMF” means soil was pasteurised before the experiment and no mycorrhizal inoculum was added.
Both plant and mycorrhizal growth were affected when the trade with mycorrhizae was high
Although, phosphorus addition stimulated photosynthesis and plant growth in both experiments, only in the first experiment did we find what we expected. When phosphorus availability was low, plants invested a greater percentage of their C budget in dealing with mycorrhizae, and less in their own growth (i.e., biomass production efficiency often called BPE, decreased) [2]. Unexpectedly in the second experiment, biomass production efficiency did not significantly change with phosphorus availability [3]. Instead, plants under lower phosphorus availability invested greater part of their C into root growth and less into aboveground respiration [3]. These contrasting results can be explained by greater C partitioning to mycorrhizae (much more “plant/mycorrhizae business” going on) in the first experiment [2]. In the second experiment, mycorrhizal abundance was so low that we were unable to measure the C allocated to it. Since mycorrhizae were not available for business – probably due to a pH decrease from 7 to 5.8 – plants needed to invest a greater percentage of their budget in root growth to increase their phosphorus range (to reach more distant shops) [3].
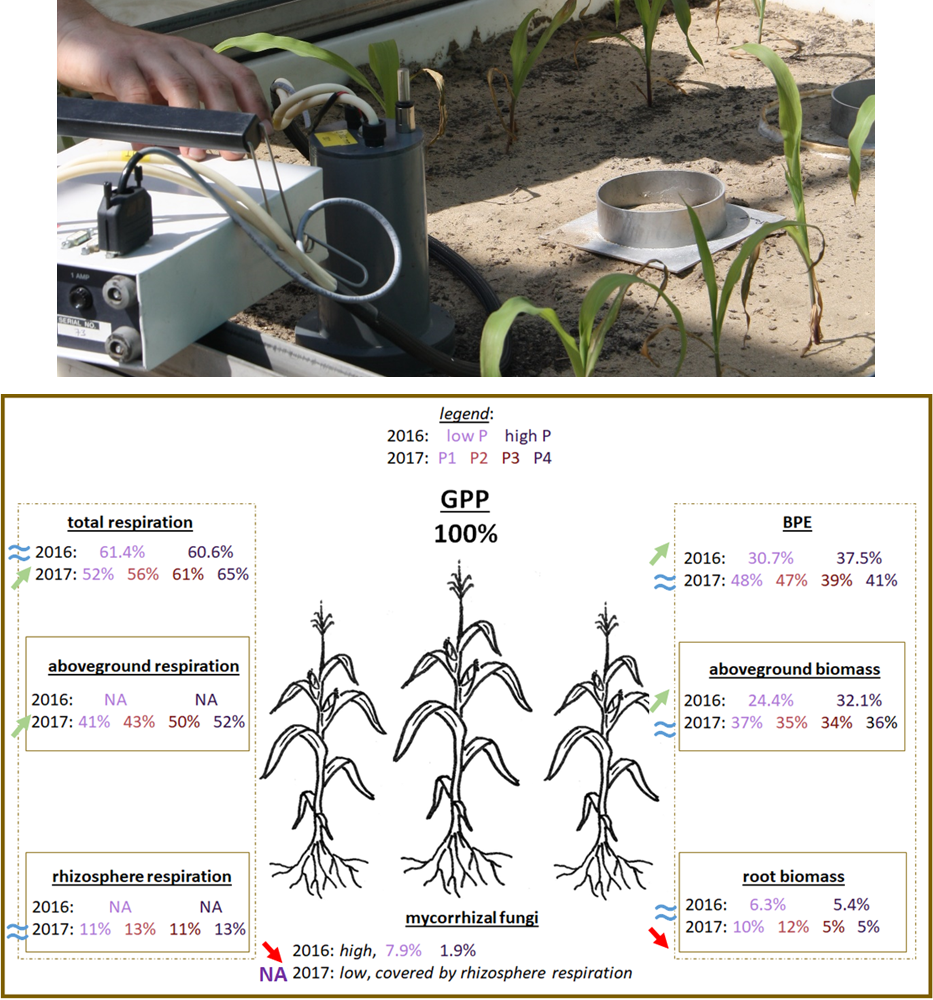
Fig. 4: top: measurement of soil respiration, bottom: C partitioning to plant biomass production (biomass production efficiency – BPE), total respiration, and mycorrhizal fungi. Results in % of total gained C (photosynthesis – gross primary production – GPP) for the 2016 experiment [2] and 2017 experiment [3]. Arrows and equality signs show significant changes in C partitioning due to changes in P fertilisation. P fertilization is low or high (2.5 kg P/ha – pink color, 20 kg P/ha – purple color, respectively) in the 2016 experiment; P1, P2, P3 or P4 (2.5, 5, 10 or 20 kg P/ha – pink, orange, red and purple color, respectively) in the 2017 experiment. Photo credit (top): Melanie Verlinden
The deal with mycorrhizae proved to be the most efficient (cheapest) way for plants to take up phosphorus in both experiments, especially in low phosphorus conditions where the plant-mycorrhizal symbiosis appeared critical for plant survival (even when mycorrhizal abundance was very low in the second experiment!) [2,3,4]. Even when phosphorus was easily available, the price that plants had to pay for phosphorus (thus the amount of C they needed to invest) was still lower when they traded with mycorrhizae than that when they grew without mycorrhizae [4]. In other words, trading nutrients with mycorrhizae came with a discount compared to direct buying at the (distant) shops. Plants used the phosphorus they purchased to grow and increase their photosynthesis (and thus increase their C budget) which more than compensated for the invested C [4].

Fig. 5 An isotopic-CO2 gas analyser was used to distinguish between soil CO2 fluxes from corn-derived rhizosphere respiration (roots, root exudates, mycorrhizae) and decomposition of (older) soil organic matter.
Being pioneers is not easy
To the best of our knowledge, this is the first experiment where the entire plant C balance has been quantified, including income, expenses, and in particular, the investment in mycorrhizae. We successfully balanced the plant’s carbon ‘budget’; plant growth matched the difference between income (photosynthesis) and other expenses (respiration and mycorrhizae). This implies that we measured the individual fluxes reasonably accurately. However, this was a major challenge. Estimating mycorrhizal abundance and C use in an experimental context is difficult since it requires a clear separation of old and new mycorrhizal biomass from other fungi and soil for analyses [5]. A critical aspect of our approach was the use of C isotopes to distinguish plant-derived C from soil C.

Fig. 6 A big cuvette connected with a CO2 analyser is enclosing a mesocosm (a miniature ecosystem) for measuring its total CO2 flux. Photo credit: Melanie Verlinden
Take-home message
Our research demonstrated how plant C partitioning is influenced by nutrient availability and mycorrhiza. We found that when there were lots of mycorrhizae, a low phosphorus availability caused plants to invest more of their C budget in mycorrhizae dealing, and less in their own growth. When there were less mycorrhizae, plants which had a low phosphorous availability invested more of their C budget in growing roots, and spent less on above ground respiration. These results emphasize the need to account not only for nutrient availability, but also for the availability and distribution of mycorrhizal symbionts when studying and modelling carbon cycling in terrestrial ecosystems.
You can find more information about our research in:
[1] Vicca, S., Luyssaert, S., Peñuelas, J., Campioli, M., Chapin, F.S., Ciais, P., … Janssens, I.A. (2012). Fertile forests produce biomass more efficiently. Ecology Letters 15: 520-526. doi: 10.1111/j.1461-0248.2012.01775.x
[2] Verlinden, M.S., Ven, A., Verbruggen, E., Janssens, I.A., Wallander, H., Vicca, S. (2018). Favorable effect of mycorrhizae on biomass production efficiency exceeds their carbon cost in a fertilization experiment, Ecology 99 (11): 2525-2534. doi: 10.1002/ecy.2502
[3] Ven, A., Verlinden, M.S., Fransen, E., Olsson, P.A., Verbruggen, E., Wallander, H., Vicca, S. (2020). Phosphorus addition increases carbon partitioning to autotrophic respiration but not to biomass production in a mesocosm experiment with Zea mays, Plant, Cell & Environment, online first. doi: 10.1111/pce.13785
[4] Ven, A., Verlinden, M.S., Verbruggen, E., Vicca, S. (2019). Experimental evidence that phosphorus fertilization and arbuscular mycorrhizal symbiosis can reduce the carbon cost of phosphorus uptake, Functional Ecology. doi: 10.1111/1365-2435.13452
[5] Ven, A., Verbruggen, E., Verlinden, M.S., Olsson, P.A., Wallander, H., Vicca, S. (2020). Mesh bags underestimated arbuscular mycorrhizal abundance but captured fertilization effects in a mesocosm experiment, Plant & Soil. doi: 10.1007/s11104-019-04368-4
Edited by Olga Vindušková
Acknowledgment:
Author and editor thank Sara Vicca, Dan Evans and Hazel Gibson for providing comments on this blog post.
Written by:

Arne Ven
Arne finished his PhD in December 2019 at University of Antwerp where he studied plant-mycorrhizal nutrient interactions under the supervision of Sara Vicca. After teaching at a local high school he is looking forward to be back at University of Antwerp this summer to work as a climate change expert for, among others, the Scientists4Climate initiative ‘Klimaatlink’ as lecturer for schools and wider public (https://www.scientists4climate.be/).
Editor’s comment: Call for more blog posts like this
Introduce your PhD research! Do your friends and family struggle to understand what is/was your PhD about? Write a blog post for them and as a side effect, promote your work among the soil science community 🙂 The SSS blog is here for you – get in touch with us!