Earth’s history is punctuated by extreme events known as mass extinctions. The End-Permian extinction, 252 million years ago, is believed to be the biggest, killing 90 % or more of all species – no wonder it is also called “The Great Dying”. The big question out there is to understand what caused it, but it is a challenge to get the complete picture of an event so long ago in prehistory. We know that the Siberian Traps (the enormous field of volcanic rock that lies in Siberia) were formed around that time and that volcanic activity was a likely trigger for this mass extinction. But what actually happened? And why did so many species, including many groups of insects, disappear?
Last year, an article appeared that brought forward a new hypothesis. From evidence in rock cores, Daniel Rothman and co-authors [1,2] concluded that around the end of the Permian, a lot of organic carbon that was formerly trapped in the sediments was converted into CO2. This change happened so fast that its release was exponential.
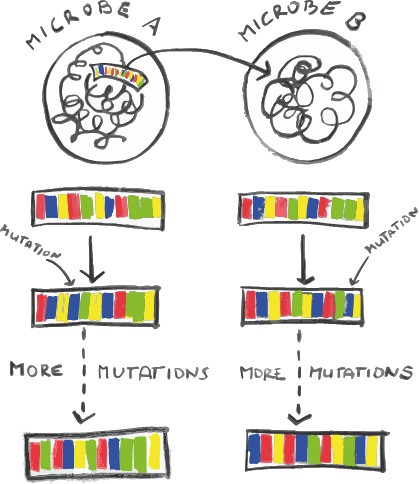
When genes are passed on from microbe A to microbe B, both undergo a certain number of mutations per period of time.
Now, what else in nature is exponential? Growth of microorganisms. So, what Rothman and his colleagues proposed is that the type of microbes that previously produced methane by using only carbon dioxide and hydrogen gas, and who had been sticking around since the very beginning of life on Earth, had suddenly acquired a new skill. They were now able to make methane out of a different source: acetate. Acetate is basically what vinegar is made out of, or what accumulates in your wine bottle when you forget about it, put the stopper on and leave it for weeks. In other words, it is the end product of anaerobic degradation. So now, armed with this new skill, the micro-organisms were able to make a lot more methane. They made so much, and rendered the atmosphere so toxic and the ocean so acidic that this caused the death of numerous land and marine organisms.
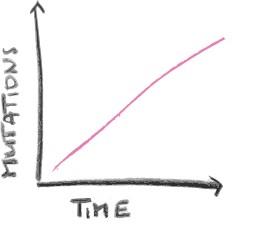
The number of differences between those two genes lets us know WHEN they started diverging, i.e. the transfer happened.
But even more recently, a new article appeared in Nature Scientific Reports investigating this topic. Here, Elliot Barnhart and co-authors presented evidence that methanogens had been using acetate for methane production for much longer (since 2.9 billion years ago)[3]. But how do we know how long ago this really was?
This is where the molecular clock comes in. Microbes can transfer genes between each other via viruses or other pathways. So, what happens quite often is that a gene gets passed on from microbe A to microbe B, in a process known as called horizontal gene transfer, as genes are transferred between individuals of the same generation. As they get passed on to their offspring (vertical gene transfer), they undergo a number of mutations at a more or less constant and known rate, which we know by looking at modern organisms. If we then compare today’s gene in microbe A and microbe B’s offspring, we can tell how many generations have passed based on the number of mutations. This tells us how long ago this transfer happened, because that was the last time both genes were identical! A mutation in this case isn’t what we might think, like growing an extra arm or ear or something – it refers to a change in the chemistry of the DNA sequence of an organism.
Why do the two different studies produce such different estimates then? Because the genes in the methanogen (Methanosarcina) that allow it to chomp away on acetate appear very similar to those of a bacterium, Clostridium, it was assumed that Clostridium had given it to the methanogen[4]. There were absolutely no genes in other methanogen-related microbes that looked remotely similar to the one allowing the use of acetate. However, the new research by Barnhart and co-authors shows that there are genes in other, methanogen-related microbes that are related to this acetate-utilization gene that had been previously overlooked. This means that methanogens could have known how to use acetate to make methane already 3.5 billion years ago, and that THEY gave it to the bacterium Clostridium around 2.52 billion years ago.
So, where are we now with the Permian-Triassic mass extinction?
Well, in addition to the causes for the Permian-Triassic mass extinction being already heavily debated for a long time anyway (there are about 130 links only on its Wikipedia page!), the authors state that methanogen activity and thus production of methane could have still exponentially increased in line with the observations made by Daniel Rothman and co-authors. The reason for this is that they need Nickel (the trace metal!) for their methane-producing enzyme to work. The volcanic activity of the Siberian Traps could have, for the first time, added so much Nickel to the biosphere that methanogens could finally rise to their full potential and “methanogenerate” the hell out of the abundantly present acetate!
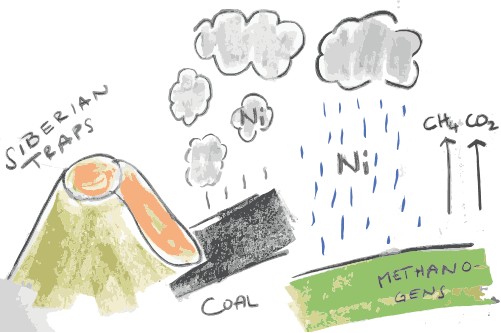
The cascade of what could have happened at the End-Permian – volcanism burns coal which releases nickel which fuels microbes.
As the authors say: “Thus, the potential for a large methane release was in place long before the end-Permian extinction, suggesting a change in environmental conditions could have promoted methanogenic microorganisms. Recent research indicates that the ancient combustion of coal through an interaction with basalt sills of the Siberian Traps released trace metals that created stimulatory conditions for methane production and instigated the end-Permian extinction [1,5–7]. Today, atmospheric methane concentrations have nearly tripled since pre-industrial times and this increase has been partly attributed to coal combustion [8,9].“ [3]
The extent of the Siberian traps, just in case you were wondering. CC BY-SA 3.0 (source)
It’s always amazing to see how little microbes play their role in the global environment, and how biochemical research can lead to new insights about their functioning in the past. Go molecular clocks!
Dr. Sabine Lengger is an organic geochemist who conducted her PhD research at the Royal Netherlands Institute for Sea Research. She has worked on archaeal lipids in sediments, contaminants in waters produced by the oil industry, on sponges and Devonian rocks, and is currently investigating lipids which can tell us more about the current and past methane cycle at the University of Bristol and lecturing Organic Chemistry at the University of Plymouth. She is into analytical chemistry, biochemical pathways, statistics, stable isotopes and likes to read about data visualization. Sabine is on Twitter (@doc_sabine), Researchgate and Google Scholar.
[1] D. H. Rothman, G. P. Fournier, K. L. French, E. J. Alm, E. A. Boyle, C. Cao, R. E. Summons. Methanogenic burst in the end-Permian carbon cycle. Proc. Natl. Acad. Sci. 2014, 111, 5462.
[2] Massachusetts Institute of Technology. Ancient whodunit may be solved: Methane-producing microbes did it! can be found under www.sciencedaily.com/releases/2014/03/140331153608.htm, 2014.
[3] E. P. Barnhart, M. A. McClure, K. Johnson, S. Cleveland, K. A. Hunt, M. W. Fields. Potential Role of Acetyl-CoA Synthetase (acs) and Malate Dehydrogenase (mae) in the Evolution of the Acetate Switch in Bacteria and Archaea. Sci. Rep. 2015, 5, 12498.
[4] G. P. Fournier, J. P. Gogarten. Evolution of Acetoclastic Methanogenesis in Methanosarcina via Horizontal Gene Transfer from Cellulolytic Clostridia. J. Bacteriol. 2008, 190, 1124.
[5] J. B. Glass, V. J. Orphan. Trace Metal Requirements for Microbial Enzymes Involved in the Production and Consumption of Methane and Nitrous Oxide. Front. Microbiol. 2012, 3, DOI 10.3389/fmicb.2012.00061.
[6] D. E. Ogden, N. H. Sleep. Explosive eruption of coal and basalt and the end-Permian mass extinction. Proc. Natl. Acad. Sci. 2012, 109, 59.
[7] M. S. Reddy, S. Basha, H. V. Joshi, B. Jha. Evaluation of the emission characteristics of trace metals from coal and fuel oil fired power plants and their fate during combustion. J. Hazard. Mater. 2005, 123, 242.
[8] P. Bousquet, P. Ciais, J. B. Miller, E. J. Dlugokencky, D. A. Hauglustaine, C. Prigent, G. R. Van der Werf, P. Peylin, E.-G. Brunke, C. Carouge, R. L. Langenfelds, J. Lathière, F. Papa, M. Ramonet, et al. Contribution of anthropogenic and natural sources to atmospheric methane variability. Nature 2006, 443, 439.
[9] S. Kirschke, P. Bousquet, P. Ciais, M. Saunois, J. G. Canadell, E. J. Dlugokencky, P. Bergamaschi, D. Bergmann, D. R. Blake, L. Bruhwiler, P. Cameron-Smith, S. Castaldi, F. Chevallier, L. Feng, et al. Three decades of global methane sources and sinks. Nat. Geosci. 2013, 6, 813.
Pingback: Morsels For The Mind – 18/09/2015 › Six Incredible Things Before Breakfast
Pingback: Morsels For The Mind – 18/09/2015 | Sandora RSS Feed Aggregator Portal