This is a guest post by Collin VanBuren. He’s currently starting his PhD at the University of Cambridge researching the effects of climate change on living and fossil frogs, as well as the relationship between the shape of anatomical structures and their function. He loves the outdoors, biomechanics, and conservation, and regularly tweets about these subjects on his Twitter account (@CollinVanBuren). He is also interested in advocating science education and one day hopes to teach and research at a university.
This is a guest post by Collin VanBuren. He’s currently starting his PhD at the University of Cambridge researching the effects of climate change on living and fossil frogs, as well as the relationship between the shape of anatomical structures and their function. He loves the outdoors, biomechanics, and conservation, and regularly tweets about these subjects on his Twitter account (@CollinVanBuren). He is also interested in advocating science education and one day hopes to teach and research at a university.
It seems that ever since the first dinosaur was mounted in a museum, palaeontologists have been curious about dinosaur limb posture and locomotion. The posture of the back legs was determined early on as being upright (completely vertical, like that of an elephant or a lion) as opposed to sprawling (splayed out to the side, like the limbs of crocodilians and most lizards). However, the jury’s still out on forelimb posture and mobility in dinosaurs.

The differences between an African elephant…

..and an american alligator are quite striking, seeing as they’re both quadrupedal animals
There are a couple of reasons why mystery surrounds this seemingly simple aspect of dinosaur biology. First, dinosaurs evolved to walk on all fours at least four times in their evolutionary history. Yep, the long-necked sauropods, horned and frilled ceratopsians, armoured thyreophorans, and ‘duck’-billed ornithopods all independently evolved quadrupedality. This transition has only occurred once outside of dinosaurs, making it extremely rare in vertebrate evolution.
The second issue is that dinosaurs had a lot of cartilage surrounding their joints. This cartilage isn’t preserved during fossilization, so anatomical information about the shoulder, elbow, and wrist joints is often lost. With these complications in mind, Matt Bonnan and I decided to try to put limitations on how the forelimb bones could have gone together in our new study in PLOS ONE.
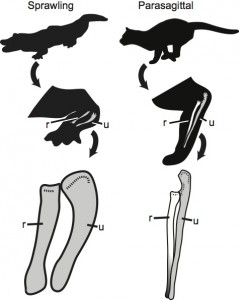
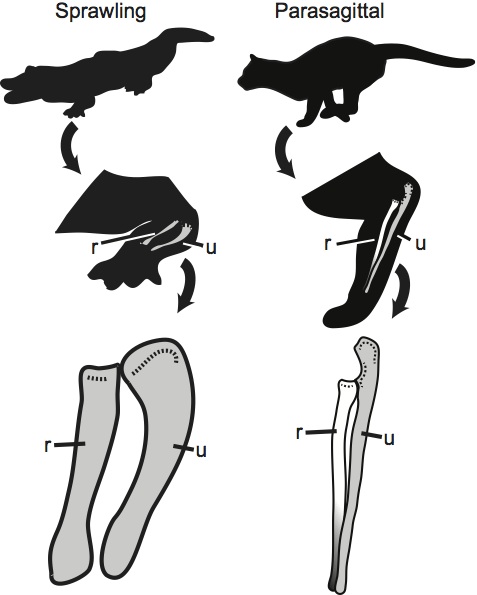
Differences in quadrupedal posture between an alligator and a cat (r – radius, u – ulna). Note the difference in how the radius crosses the ulna.
For this study, we focused on the radius bone, one of the bones of the forearm (the area between the elbow and the wrist). In living animals the curvature of the radius and the shape of the radial head determine if the hand and wrist can rotate. This motion comes in handy when we open doors or screw in light bulbs, for example. The results of our study showed that dinosaurs had a limited or non-existent ability to do this motion, at least in the way that we mammals can (some lizards have been found to do this motion in a different way, but without the same range of motion).
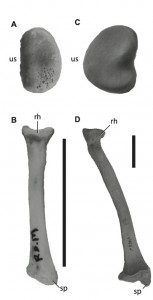
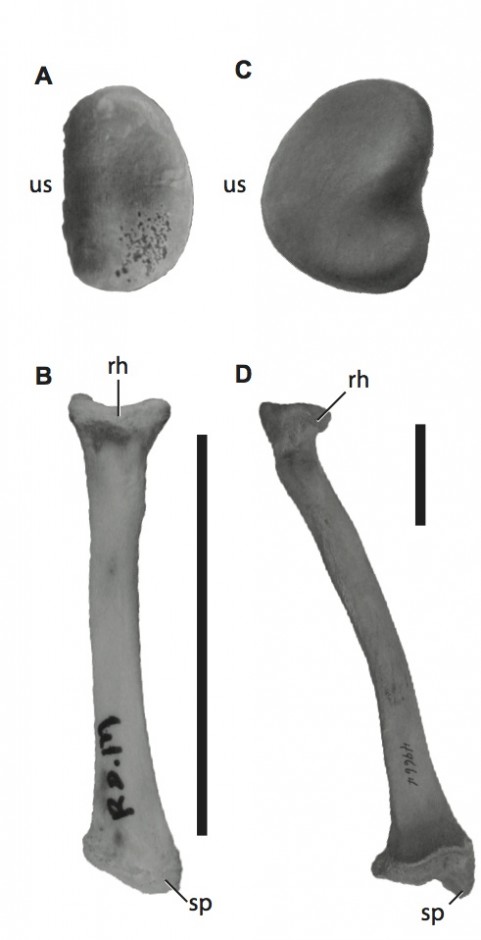
The radial head (A) and long axis (B) of Caiman crocodylus and the radial head (C) and long axis (D) of Ursus americanas. Scale bar = 5 cm. Radial heads not to scale. rh, radial head; sp, styloid process; us, ulnar articular surface.
Having a curved radius is also how a lot of mammals obtain a upright forelimb posture. Mammals that don’t have a curved radius usually have a large radius and significantly reduced ulna (the other forearm bone; horses and other hoofed mammals do this). Dinosaurs had radii and ulnae of roughly equal proportion and have straight radii like crocodilians and most lizards, meaning that, if quadrupedal dinosaurs did have upright forelimbs, they achieved this posture in a way we haven’t seen in anything alive today. In fact, other recent studies on the hands of dinosaurs have found results that seem to support our conclusion of potentially ‘weird forelimbs in dinosaurs.’
What does this all mean for how dinosaurs walked their walk? We now know that dinosaurs didn’t have the ability to turn doorknobs or screw in light bulbs, but future studies on dinosaur locomotion that take our limitations into account will have to be performed before we know for sure. The mystery of dinosaur forelimb posture isn’t completely solved yet, but hopefully we’re one ‘step’ closer to reaching that goal.
Article: http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0074842 (open access!)